У грибов различают два типа размножения: вегетативное и репродуктивное.
Вегетативное размножение осуществляется частями вегетативного тела или спорами, которые образуются клетками вегетативных гиф (рис. 16). Самая примитивная форма вегетативного размножения - частицами гиф (а также обрывками шнуров, ризоморф, склероциями), которые, будучи отделены от материнского мицелия и попав в благоприятную среду, могут дать начало новому самостоятельному мицелию.
У некоторых грибов, например дрожжевых, вегетативное размножение осуществляется почкованием клеток. На определенном этапе развития на них появляются небольшие выросты, которые постепенно увеличиваются в размерах, затем отделяются от материнской клетки и в свою очередь начинают почковаться. Такие клетки называют бластоспорами.
Своеобразная форма вегетативного размножения - образование оидий и хламидоспор. Оидии образуются в результате распадения гиф, начиная с их концов, на отдельные членики, которые в дальнейшем дают начало новому мицелию. Оидии снабжены тонкой оболочкой и недолговечны. Они встречаются у голосумчатых и других грибов. Разновидностью оидий являются геммы, отличающиеся от них более плотной темной оболочкой и способностью дольше сохраняться (например, в зимний период). Геммы известны у многих сумчатых, несовершенных, а также некоторых головневых грибов.
Хламидоспоры возникают путем уплотнения и обособления содержимого отдельных клеток мицелия, которое при этом покрывается толстой темноокрашенной оболочкой. Освободившиеся из клеток материнских гиф хламидоспоры способны долго сохраняться при неблагоприятных условиях. Прорастая, они образуют органы спороношения или мицелий. Хламидоспоры образуются у многих базидиомицетов (например, головневых), дейтеромицетов, оомицетов.
Репродуктивное размножение происходит с помощью спор, которые образуются внутри или на поверхности специальных органов, отличающихся по своему строению от вегетативных гиф. Репродуктивное размножение может быть бесполым - с образованием спор без оплодотворения, и половым, при котором образованию спор предшествует половой процесс.
Бесполое размножение. Как правило, бесполое спороношение грибов формируется на хорошо развитом мицелии, имеющем достаточный для образования спор запас питательных веществ. Спороносными органами при бесполом размножении грибов служат специальные ответвления или участки гиф, внутри или снаружи которых образуются споры (рис. 17).
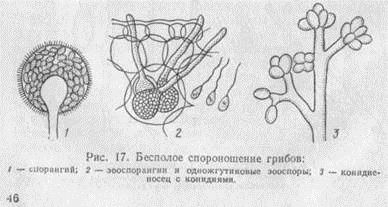
Простейшая форма бесполого размножения низших грибов - образование зооспорангиев, представляющих собой расширенные окончания гиф. Их содержимое распадается на подвижные споры с одним или двумя жгутиками - зооспоры, которые могут передвигаться и сохранять жизнеспособность только в воде. Поэтому зооспоры свойственны водным и некоторым почвенным грибам, а у наземных видов, сохранивших связь с водной средой, образуются во время дождя, росы или густого тумана.
Другой формой бесполого размножения низших грибов являются спорангии - шаровидные вместилища на концах ответвлений мицелия. Ветвь, несущая спорангий, называется спорангиеносцем. Внутри спорангия формируются неподвижные одноклеточные споры - спорангиоспоры. Они одеты твердой оболочкой; образуются у мукоровых грибов.
Наиболее распространенная форма бесполого размножения, свойственная почти всем высшим и некоторым низшим грибам,- конидиальное спороношение. Конидиями называют споры, образующиеся на конечных ответвлениях специальных органов - конидиеносцев. Конидиеносцы и конидии очень разнообразны по форме, размерам, строению и окраске, а также по характеру их развития и размещения. Разновидностями конидиального спороношения грибов являются коремии, ложа и пикниды.
Коремии - это пучки плотно соединенных друг с другом конидиеносцев, на концах которых образуются конидии. Скопления конидий на коремиях могут быть порошащими или слизистыми. Спороношения коремального типа часто встречаются у грибов из порядка гифомицетов, у сумчатых грибов.
Ложа представляют собой подушковидные, более или менее плотные мицелиальные образования, формирующиеся на поверхности пораженного субстрата или погруженные в него, но при созревании выступающие наружу. Конидиеносцы располагаются на поверхности ложа сплошным слоем. Такая форма конидиального спороношения характерна для грибов порядка меланкониевых.
Пикниды - это шаровидные (иногда немного приплюснутые) или грушевидные вместилища с плотной, обычно темной оболочкой. Они бывают одиночными (иногда их называют однокамерными) или образуются группами в общей строме (многокамерные пикниды). Конидии в пикнидах, или пикноспоры, образуются на коротких конидиеносцах, покрывающих внутреннюю стенку пикниды. Созревая, они выделяются из пикниды через специальное отверстие (устьице) сплошной слизистой массой, застывающей на воздухе в виде капель или скрученных нитей, нередко имеющих яркую окраску. Чаще всего пикниды частично или полностью погружены в питающий субстрат и наружу выступают лишь их верхние части или устьица. Спороношение типа пикнид - основной признак грибов порядка пикнидиальных.

У многих фитопатогенных грибов в течение вегетационного периода наблюдается несколько генераций бесполого спороношения; этим обеспечиваются массовое распространение инфекционного начала и повторное заражение растений.
Половое размножение. Половой процесс у грибов весьма разнообразен (рис. 18). В простейшей форме он представлен изогамией, т. е. слиянием двух одинаковых по форме, но разнополых зооспор. В результате формируется циста. У некоторых наиболее примитивных грибов (например, слизевиков) продуктом полового процесса типа изогамии является диплоидный амебоид.
Более сложные формы полового размножения - оогамия и зигогамия. При оогамии на мицелии закладываются различные по форме и строению половые клетки: женская - оогоний и мужская - антеридий. После слияния их содержимого образуется ооспора. При зигогамии сливается содержимое двух внешне одинаковых клеток разнополых мицелиев. В результате образуется зигоспора. Цисты, ооспоры и зигоспоры - это покоящиеся споры, покрытые толстой оболочкой и предназначенные для сохранения вида при неблагоприятных условиях. Они характерны для низших грибов.
У высших грибов (сумчатых и базидиальных) половое воспроизведение завершается формированием сумок или базидий. Сумки (аски) - это различной формы мешковидные клетки, внутри которых развиваются сумкоспоры (аскоспоры). Обычно сумка содержит восемь сумкоспор. Базидия представляет собой тело булавовидной или цилиндрической формы, на поверхности которого образуются базидиоспоры. Чаще всего их четыре.
Грибы, у которых мицелий наделен свойствами обоих полов и половой процесс осуществляется путем слияния цитоплазмы и ядер клеток одного и того же мицелия, называются гомоталличными. Грибы, у которых каждый мицелий обладает свойствами только одного пола, а в половом процессе участвуют клетки и ядра разнополых мицелиев, называются гетероталличными. В связи с тем что мицелии разного пола у гетероталличных грибов морфологически неразличимы (у многих видов нет четких морфологических различий даже между копулирующими клетками), конкретный пол у грибов часто не указывают и половую принадлежность отдельных особей обозначают знаками плюс (+) и минус (-).
Переход к половому размножению у грибов часто бывает связан с окончанием вегетативного роста, наступлением неблагоприятных условий, перезимовкой. У фитопатогенных видов половое спороношение, как правило, наблюдается один раз в течение года, а споры, образующиеся половым путем, осуществляют первичное заражение растений весной или в начале лета.
Способность грибов формировать в цикле развития разные типы спороношений называется плеоморфизмом. Циклы развития большинства грибов включают два спороношения - половое и бесполое, которые чаще всего имеют самостоятельные видовые названия. В то же время существует немало видов, имеющих наряду с половым несколько различных бесполых спороношений. Такой цикл развития называют сложным. Примером грибов со сложным циклом развития могут служить ржавчинные грибы. Дейтеромицеты (несовершенные грибы) имеют только бесполые спороношения. У некоторых сумчатых и базидиальных грибов известно лишь половое спороношение.
Споры грибов, предназначенные для распространения и быстрого прорастания в течение вегетационного периода, называются пропагативными. Такие споры имеют тонкие, обычно бесцветные оболочки и не могут долго сохранять жизнеспособность. Споры, служащие для длительного сохранения при неблагоприятных условиях, называются покоящимися. Они бывают снабжены толстой и, как правило, пигментированной оболочкой.
Спорообразующая способность большинства грибов очень велика. Например, у мукоровых грибов один спорангий может содержать несколько десятков тысяч спорангиоспор. В каждой пустуле ржавчинных грибов созревает несколько сотен спор. Число сумкоспор в плодовом теле некоторых сумчатых грибов исчисляется миллионами, а плодовые тела многих трутовых грибов выделяют миллиарды базидиоспор ежедневно в течение всего периода споруляции. Количество продуцируемых грибами спор многократно увеличивается соответственно числу генераций бесполого спороношения.
Вегетативным называется такое размножение, которое осуществляется неспециализированными или малоспециализированными частями вегетативного тела, т. е. мицелия гриба.
Оно осуществляется при помощи частиц или обрывков живой грибницы, которые, расселяясь различными путями, могут дать начало новому мицелию. Этот наиболее простой способ вегетативного размножения грибов очень распространен в естественной обстановке и практикуется при искусственном разведении шампиньонов, а также при пересевах чистых культур грибов в лабораторных условиях.
Вегетативное размножение возможно отдельными кусочками склероция, ризоморфы и мицелиального тяжа, которые, попадая в благоприятные условия, прорастают и дают начало новому грибу. Такое распространение типично для домового гриба, опенка и других грибов. Более специализирован способ вегетативного размножения, при котором грибница благодаря особенностям своего строения легко разъединяется на отдельные клетки или споры, каждая из которых в дальнейшем вырастает в новую такую грибницу. К таким спорам вегетативного размножения относятся хламидоспоры, оидии, геммы и другие видоизменения мицелия.
Хламидоспорами называют участки гиф, обособившиеся от вегетативного мицелия и покрытые плотной утолщенной и иногда пигментированной оболочкой. Они содержат значительные запасы питательных веществ, а благодаря плотной оболочке способны длительно (до 10 лет и больше) сохраняться, перенося неблагоприятные условия (сухость, низкие или высокие температуры и пр.).
Оидиями называют короткие округлые или удлиненные членики грибницы, покрытые тонкой оболочкой; долго сохраняться не могут, но при прорастании дают начало новой грибнице.
Геммы по способу образования напоминают хламидоспоры, отличаясь от них непостоянством формы. Они известны у большого числа сумчатых и несовершенных грибов в качестве органа перезимовки.
Бесполое размножение осуществляется посредством особых спор, развивающихся на специальных резко обособленных ветвях мицелия. По происхождению споры могут быть эндогенными и экзогенными. Эндогенные споры образуются обычно в большом числе в особых клетках - спорангиях, почему сами споры называются спорангиоспорами . Экзогенные споры образуются на поверхности производящих их органов, называемых конидиеносцами , а сами споры - конидиями. У низших грибов преобладает образование спорангиоспор, у немногих встречаются конидии, у высших грибов имеются только конидии.
Спорангии образуются на особых ветвях грибницы - спорангиеносцах, отличающихся большей толщиной, ограниченным ростом и другими признаками. Примером такого способа бесполого размножения может быть головчатая плесень (Mucor alboater N. N.), развивающаяся иногда на ослабленных семенах сосны, ели и других хвойных при их проращивании, а также Rhizopus nigricans Ehrenb. на плодах грецкого ореха, самшита и других лиственных пород.
У более примитивных форм типом бесполого размножения является образование зооспорангия, отличающегося от спорангия тем, что содержимое его распадается на подвижные споры, снабженные одним или двумя жгутиками и получившие название зооспор. Примером такого способа размножения может быть Phytophthora omnivora DB., поражающая всходы хвойных, бука и других древесных пород.
Другой тип бесполого размножения у грибов - образование конидий, развивающихся на конидиеносцах различной степени сложности. Они могут быть одиночные и неветвящиеся, или сильно ветвящиеся и несущие на концах ветвей группы конидий. Конидия закладывается всегда как одна клетка, имеющая округлую или овальную форму. У многих несовершенных грибов конидия так и остается одноклеточной. В некоторых случаях она изменяет свою форму, приобретает поперечные и даже продольные перегородки и в завершенном виде представляет сложное многоклеточное тело, как например, Alternaria coryli Israf. на мелких орехах при их хранении или Alternaria tenuis Nees на желудях дуба.
Коремии представляют собой тесно сближенные и спаянные вдоль конидиеносцы, отшнуровывающие на своих концах конидии. Они характерны для целого порядка несовершенных грибов, называемого Coremiales. Его представителем может быть Graphium ulmi Schw. - опасный и наиболее распространенный возбудитель сосудистого заболевания вяза - голландской болезни.
Ложе состоит из плотного сплетения гиф мицелия, на поверхности которого располагаются короткие конидиеносцы с конидиями. Часто ложа закладываются внутри ткани питающего растения и долгое время остаются прикрытыми, освобождаясь лишь через разрыв наружных покровов к моменту созревания конидий. В результате на пораженных частях растений появляются резко очерченные пятна или язвы, которые характерны для заболевания, называемого антракнозом . Конидиальные спороношения в форме ложа типичны для некоторых несовершенных грибов, объединяемых по этому признаку в порядок меланкониевых (Melanconiales) с наиболее распространенными родами Gloeosporium, Colletotrichum и др.
Пикниды представляют собой шарообразные полые вместилища, наиболее часто с бурой оболочкой, состоящей из сплетения грибницы пара плектенхиматического и прозоплектенхиматического строения, с узким отверстием - устьем наверху. Внутри полости пикниды находятся тесно скученные короткие конидиеносцы, отделяющие внутрь пикниды конидии, иногда называемые пикноспорами , или стилоспорами .
Пикниды могут быть одиночными или соединенными по нескольку в общей строме, имеющей одно общее устье. Пикниды характерны для большой группы несовершенных грибов порядка Pycnidiales, например, Ascochyta velata Kab. et Bub. - на листьях и крылатках клена остролистного, Phyllosticta platanoides Sacc. - на крылатках семян клена остролистного, клена татарского, Septoria quercina Desmaz. - на листьях дуба (серая пятнистость), Septoria salicicola Sacc. - на листьях ивы.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Грибы размножаются спорами , которые представляют собой одноклеточное образование, с помощью которого и осуществляется бесполое размножение . Спору можно сравнить с очень просто устроенным семенем , не имеющим зародыша и представляющим комочек протоплазмы, защищенный плотной оболочкой .
Грибы размножаются спорами
Количество спор гриба
Можно представить, какое огромное потомство способно произвести лишь одно плодовое тело гриба!
Прорастание спор
На самом деле это не происходит, так как прорастает спора
лишь тогда, когда она попадет в условия, благоприятные для ее развития, а это происходит редко. Объясняется это особыми условиями, необходимыми для жизни разных грибов.
Обычное в природе явление: чем меньше у спор шансов оказаться в условиях, благоприятных для прорастания, тем их больше.
Спорообразование у грибов чередуется с половым размножением, в общем сходным с этим процессом у водорослей.
 Процесс размножения водорослей сходный с грибами
Процесс размножения водорослей сходный с грибами
Кроме размножения с помощью спор, грибы размножаются и другими способами:
- дрожжевые грибки - особыми «почками»,
- многие другие - кусочками грибницы. Так, например, искусственно размножают вкусный шляпочный гриб .
Форма плодовых тел может быть очень разнообразной. У большинства шляпочных грибов в плодовом теле различают пенек, или ножку, и шляпку на ней. Но плодовое тело может иметь и иную форму (дождевики, трюфели, ).
Споры шляпочных грибов
Споры у шляпочных грибов развиваются под шляпкой, в тесно прижатых друг к другу трубочках (губчатые грибы - белый, масленок, подосиновик и другие), или на особых пластинках, расположенных снизу шляпки (пластинчатые грибы- , и другие).
 Споры у шляпочного гриба рыжика развиваются на особых пластинках, расположенных снизу шляпки
Споры у шляпочного гриба рыжика развиваются на особых пластинках, расположенных снизу шляпки
Есть грибы, у которых споры образуются на особых сосочках, под шляпкой (желтый и серый колчаки), на ее верхней поверхности в особых извилинах - складках (сморчки, строчки), внутри плодового тела (дождевики, трюфели).
Если шляпку зрелого пластинчатого гриба положить на лист бумаги, а затем, спустя некоторое время, осторожно снять ее, то на бумаге обозначатся радиальные линии, идущие во все стороны от центра пенька. Это выпавшие из пластинок споры.
Споры эти имеют разную окраску - у шампиньона они темные, а у обыкновенного красного мухомора - белые. Поэтому подкладывать бумагу для обнаружения спор нужно не одного с ними цвета.
По форме и окраске спор возможно точно определить виды грибов. Это особенно важно в случаях отравления . Микроскопическое исследование дает возможность определить вид гриба по спорам, оказавшимся в пище.
Мухомор — ядовитый грибСпоры многих грибов выдерживают высушивание и низкие температуры, не теряя способности прорастать. Их много носится в воздухе вместе с пылью.
Пылинка-спора, попав в благоприятные для жизни гриба условия, образует маленький вырост - гифу , которая, разрастаясь, дает грибницу. Но эти условия, необходимые для прорастания споры, у многих шляпочных грибов нередко очень сложны.
Так, учеными установлено, что споры некоторых грибов становятся способными прорастать лишь в том случае, если пройдут через кишечник личинок некоторых насекомых, живущих на этих грибах. «Червяки», которыми иногда переполнена мякоть гриба, и являются личинками разных насекомых - «грибных мушек», откладывающих яйца в плодовое тело.
Бесполое размножение реализуется путём образования конидий и спорангиоспор, содержащих весь генетический материал, необходимый для возникновения и развития новой колонии. Если у гриба неизвестны половые формы, то строение конидий используют как классифицирующий признак. Грибы, не размножающиеся половым путём, или виды, для которых совершенные (половые) формы неизвестны, объединены в класс Deuteromycetes, или Fungi imperfecti (несовершенные грибы).
Конидии - неполовые репродуктивные структуры. Они могут быть одноклеточными (микроконидии) либо многоклеточными (макроконидии). У некоторых грибов конидии формируются на концах конидионосных гиф- конидиофоров (бластоконидии), у других- по бокам и на концах обычных гиф (таллоконидии) путём их септирования и расчленения. Бластоконидии (бластоспоры) формируются в результате отпочковывания от конидиофора. Их образование характерно для дрожжей и дрожжеподобных грибов. Наиболее типичные таллоконидии - артро- и хламидоконидии.
Артроконидии (артроспоры ) образуются в результате фрагментации септированных гиф, распадающихся на отдельные клетки (рис. 2-10). Хламидоконидии (хламидоспоры) - таллоконидии, образующиеся внутри нитей мицелия -в виде увеличенных клеток, окружённых толстой оболочкой. За счёт плотной оболочки хламидоконидии приобретают устойчивость к различным воздействиям и сохраняются после гибели и распада мицелия (отдалённо напоминая этим цисты простейших). Выделяют хламидоконидии терминальные (концевые), располагающиеся на конце гиф, и интеркалатные (промежуточные), располагающиеся в середине гифы.
Рис. 2-11. Бесполое размножение грибов с образованием фиалоконидии .
Спорангиоспоры - споры , образующиеся вегетативным путём в специализированных мешках (спорангиях). Последние располагаются на концах особых гиф - спорангиофорах. К основным типам спорангиев относят фиалиды {образования, напоминающие бокал), содержащие фиалоспоры, или фиалоконидии (рис. 2-11) и аннелиды (кольцевидные структуры), содержащие аннелоспоры, или аннелоконидии.
Бесполое спороношение грибов. Наиболее примитивным органом бесполого размножения у низших грибов является зооспорангий, представляющий собой расширенное окончание гифы, внутри которого формируются подвижные споры с одним или двумя жгутиками – зооспоры.Более совершенной формой бесполого размножения низших грибов является образование спорангиев – шаровидных вместилищ, на концах ответвлений грибницы. Ветвь, несущая спорангий, называется сиорангиеносцем. Внутри спорангия формируются неподвижные споры – спорангиоспоры. Самой распространенной формой бесполого размножения, свойственной высшим грибам, является конидиальное спороношение. Конидии – это споры, образующиеся на концах вегетативных гиф или на конечных ответвлениях специальных органов – конидиеносцев.Конидиеносцы и конидии очень разнообразны по форме, размерам, строению и окраске, а также по характеру развития и размещения.Бесполое спороношение у фитопатогенных грибов обычно образуется многократно в течение вегетационного периода и служит для массового распространения гриба и повторного заражения растений.
Половое размножение грибов
Среди грибов, патогенных для человека, способность размножаться половым путём выявлена у представителей классов Ascomycetes, Basi-diomycetes и Zygomycetes. Половым путём образуются аскоспоры, базидиоспоры и зигоспоры.Аскоспоры. У аскомицетов половым путём в специализированных булавовидных (реже шаровидных) клетках - асках - образуются споры, число которых всегда кратно двум.

Базидиоспоры образуются после мейотического деления спор на базидии - концевой клетке мицелия. Обычно на базидии развивается четыре споры.Зигоспоры . Зигомицеты могут размножаться как вегетативным, так и половым путём . При половом размножении верхушки близко расположенных гиф соединяются, их оболочки растворяются, ядра сливаются (как и у прочих эукариотов). Процесс проходит постадийно: первоначально сливаются два протопласта (плазмогамия), затем оба ядра сопряжённо делятся и после образования плодового тела сливаются (кариогамия). За кариогамией следует мейоз - редукционное деление, при котором число хромосом уменьшается до исходного гаплоидного. Затем образуется зигоспора с толстыми ригидными стенками. Процесс образования зигоспор известен как зигогамия .

половое размножение у грибов бывает различных типов. Сущность его заключается в том, что происходит слияние двух половых клеток (гамет) - мужской и женской - или двух вегетативных талломов, функционирующих как половые клетки, в результате возникает новообразование (зигота). Сливающиеся гаметы содержат только половинный набор хромосом. В зиготе число хромосом соответственно удваивается. Гаметы являются структурами, которые находятся, имея половинный хромосом, в гаплоидной фазе, а зигота переходит уже в диплоидную фазу.У высших грибов половой процесс протекает как слияние органов и клеток, не дифференцированных на гаметы. Образовавшаяся в результате слияния зигота (также не дифференцированная и обычно представляющая собой лишь соответствующее ядерное состояние) без периода покоя переходит к дальнейшему развитию; в ней формируются дикарионы ядер противоположных полов, которые потом попарно сливаются и претерпевают редукционное деление. Гаплоидные ядра, которые образовались в процессе редукционного деления, переходят в аскоспоры, образующиеся в сумках или в базидиоспоры, образующиеся на специальных клетках - базидиях - базидиомицетах экзогенно.
16. Споры грибов, развивающиеся в период спороношения, могут отличаться. Одни споры - пропагативные - развиваются в огромном количестве, служат для быстрого расселения грибов. Так, в плодовом теле дождевика - до 7,5 миллионов спор, а шампиньон за 5 дней рассеивает более 10 биллионов спор. Такие споры быстро прорастают, но являются маложивучими, часто недозрелыми и в больших количествах погибают. Другие споры - покоящиеся - развиваются в малом количестве, нуждаются в покое, во время которого происходит их дозревание, служат для сохранения вида при неблагоприятных условиях. Такие споры могут сохраняться до 10-12 лет и прорастают медленно. Прорастают они обычно спорангием, дающим массу спор.
17. Царство грибов характеризуется разнообразием жизненных циклов и вариантов ядерного статуса (плоидность, количество ядер в клетке, их генетическая разнородность или однородность). У различных таксономических групп грибов выделяют до 7 типов жизненного цикла (по Дж. Рэйперу ).
1. Бесполый цикл характерен для нескольких десятков тысяч видов аскомицетов ибазидиомицетов, утративших половую стадию - так называемых дейтеромицетов.
2. Гаплоидный цикл известен у зигомицетов, многих хитридиомицетов. Мицелий содержит множество гаплоидных ядер (мультикариотический мицелий), которые делятся митотически по мере роста гиф.
3. Гаплоидный цикл с ограниченным дикарионом характерен для большинства аскомицетов, мицелий их также чаще всего бывает гаплоидный мультикариотический. Гаметы или гаметангии вначале сливаются цитоплазмами (происходит плазмогамия ) без слияния ядер (кариогамии ) и прорастают дикариотическими гифами, называемыми такжеаскогенными .
4. Гаплоидно-дикариотический (гапло-дикариотический) цикл встречается у многих базидиомицетов - гименомицетов,гастеромицетов, ржавчинных грибов. Он сходен с предыдущим, но характеризуется длительной стадией дикариона, которая чаще всего бывает доминирующей.
5. Дикариотический цикл характерен для ограниченной группы базидиомицетов - головнёвых грибов. Гаплоидная фаза у них представлена базидиоспорами и прорастающими из них одноядерными споридиями , которые способны расти на питательной среде. Сливаясь попарно, споридии формируют дикариотический мицелий.
6. Гаплоидно-диплоидный (гапло-диплоидный) цикл встречается у низших водных грибов - бластокладиевых, хитридиомицетов. Диплоидный спорофит образует зооспоры, прорастающие в такие же диплоидные спорофиты (бесполое размножение) и мейоспорангии, дающие начало гаплоидному половому поколению - гаметофитам..
7. Диплоидный цикл известен у дрожжей Saccharomyces cerevisiae и других сахаромицетов, кроме настоящих грибов характерен также для оомицетов, например, Phytophthora infestans . Преобладает диплоидная стадия почкующихся клеток, ядра которых в определённых условиях (например, истощение питательной среды) делятся мейозом и формируют гаплоидные аскоспоры..
Часто один и тот же гриб образует в цикле своего развития не одно, а несколько спороношений. Они чередуются, следуя одно за другим, и каждый раз гриб меняет форму. У некоторых грибов и ветегативное тело может видоизменяться на протяжении жизни, переходя в зависимости от условий внешней среды от мицелиального роста к дрожжеподобному и наоборот. Такие изменения формы грибов в течение жизни получили название полиморфизма. Полиморфизм, связанный со сменой дрожжевой и мицелиальной форм роста в ходе вегетативного развития гриба обычно называют диморфизмом, а со сменой разных типов спороношений - плеоморфизмом. Явление плеоморфизма грибов вызывает большие затруднения в работе микологов. Действительно, если не исследовать связи между отдельными спороношениями, каждое из них можно принять за самостоятельный гриб
ПЛЕОМОРФИЗМ- своеобразный тип полиморфизма, проявляющийся в способности одного вида гриба образовывать разные спороношения на различных стадиях своего развития. В жизненном цикле развития этих грибов наблюдается последовательная смена таких спороношений, которые в большинстве случаев резко отличаются друг от друга по морфологии. П. известен у сумчатых, ржавчинных и некоторых др. групп грибов.
18. КЛАСС СУМЧАТЫЕ ГРИБЫ ИЛИ АСКОМИЦЕТЫ (ASCOMYCETES)
Сумчатые грибы, или аскомицеты,- один из крупнейших классов грибов. В нем более 30 000 видов, что составляет около 30% всех известных видов грибов. Входящие в этот класс грибы чрезвычайно разнообразны по строению. Сюда относятся, например, дрожжи, представленные одиночными почкующимися клетками, и виды с плодовыми телами различной формы и размеров, от микроскопических до крупных, достигающих иногда 10-20 см (сморчки, строчки).
У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших - в специальных вместилищах - плодовых телах (рис. 69). Различают следующие типы плодовых тел: клейстотеций (полностью замкнутое плодовое тело), перитеций (полузамкнутое, обычно кувшиновидпое плодовое тело с отверстием на вершине) и апотеций (открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз). Клейстотеций, перитеции и апотеции - настоящие плодовые тела, развитие которых происходит по аскогимениальному типу - образование их оболочки (перидия) происходит одновременно с развитием аскогенных гиф и сумок.Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах - плотных сплетениях гиф различной формы, размера и консистенции. Однако они всегда имеют собственный перидий, заметный хотя бы на ранних стадиях развития плодового тела. 
19. Плодовые тела базидиомицетов различны по форме и консистенции. Они могут быть паутинистыми, рыхлыми, плотно-войлочными, кожистыми, деревянистыми, мягкомясистыми, могут иметь форму пленок, корочек, могут быть копытообразными или состоять из шляпки и ножки.
Спороносный слой плодового тела - гимений - располагается у более примитивных видов на верхней стороне плодовых тел, а у более высокоорганизованных - на нижней. Гимений базидиальных грибов состоит из базидий с базидиоспорами и парафиз. У некоторых видов в гимении находятся цистиды - крупные клетки, возвышающиеся над гимениальным слоем. Они защищают гимениальный слой и особенно базидий от давления сверху.
Поверхность плодового тела, несущую гимении, называют гименофором. У низших представителей он гладкий, а у более высокоорганизованных имеет форму зубцов, трубочек, пластинок (рис. 155, 156, 157).

ХИТРИДИОМИЦЕТЫ
Половое размножение представлено своеобразной оогамией. Оогоний содержит одну или несколько яйцеклеток, в то время как антеридий не образует обособленных гамет и приоплодотворении просто переливает часть своего клеточного содержимого с ядрами в оогоний, что является характерным признаком, по которому выделена группа. Образующиеся после оплодотворения ооспоры являются покоящейся формой оомицета, в которой он пережидает неблагоприятный период, например, зиму. Весной они прорастают, давая зооспорангий. Таким образом, ооспора к тому же и единственная диплоидная стадия в цикле развития оомицетов. Для оомицетов характерен гинадромиксис (от «гина» - женский, «андрос» - мужской), т.е. смена пола при наличии определенных феромонов (антеридиол, оогониол). Встречается только у двудомных, у которых антеридии и архегонии развиваются на разных талломах.Классификационное положение
Несмотря на наличие развитого мицелия, оомицеты были исключены из числа грибов. Из всех грибов лишь они и хитридиомицеты имеют подвижные зооспоры, оомицеты не имеют хитина в клеточной стенке, биосинтез лизина происходит у них как у растений, а не как у остальных грибов, наконец, они имеют совершенно не свойственную грибам ультраструктуру митохондрий. На основании этих признаков в конце XX векацарство грибов было разделено на два отдела: Oomycota и Eumycota , в наше время отдел Oomycota с классом Oomycetes перенесён в царство Chromista ; по другой классификационной схеме класс Oomycetes включается в тип Heterokontophyta царства Protista .
Из группы пероноспоровых в Ленинградской области особенно распространена фитофтора инфестанс, вызывающая фитофтороз картофеля. Гриб поражает и листья, и клубни картофеля. На зараженных листьях образуются крупные бурого цвета пятна, развивающиеся от края листовой пластинки к центру. На границе между больной и здоровой тканью хорошо заметен с нижней стороны листа нежный белый налет. Он состоит из выступающих из устьиц спорангиеносцев, отчленяющих лимоновидные зооспорангии, которые подхватывает ветер и переносит на новые здоровые растения. Кроме картофеля гриб поражает томаты, развиваясь как на листьях, так и на плодах. известной под названием ложной мучнистой росы или пероноспороза. возбудители ложной мучнистой росы капусты (пероноспора капустная), клевера (пероноспора клеверная), лука (пероноспора Шлейдена) и многие другие. Наиболее опасным является пероноспороз капусты, гороха и клевера. Пероноспороз капусты, например, проявляется в течение всей вегетации, но особенно он опасен при поражении рассады и семенников. Пероноспороз гороха и клевера в периоды массовых вспышек заболевания (эпифитотии) может нанести большой ущерб как молодым растениям, так и семенникам. В последнем случае семян чаще всего не образуется, а если они и образуются, то имеют небольшой процент всхожести. На поверхности пораженных органов развиваются конидии. С их помощью гриб распространяется и, попадая на листья нового растения, заражает его.
22. Пероноспоровые по сравнению с сапролегниевыми и лептомитовыми представляют собой следующий этап приспособления грибов к жизни на суше. Это довольно древняя группа, возникновение которой относят к палеозойской эре (карбон), наиболее эволюционировавшая среди ооцицетов. В ней насчитывается несколько сотен видов, причем наибольшее их число входит в семейство пероноспоровых (Peronosporaceae), представители которого характеризуются более совершенными признаками, тогда как другие семейства содержат значительно меньше видов и эволюционно менее прогрессировали.Порядок пероноспоровых делят на 3 семейства: питиевые, цистоповые и пероноспоровые.По приуроченности к среде Обитания виды пероноспоровых грибов можно отнести к гидрофилам, зоофилам и фитофилам.По характеру развития пероноспоровые грибы - эфемеры. Они требуют короткого времени для образования спороношения и затем относительно быстро отмирают, но при помощи своих спор, попавших на благоприятные субстраты, развиваются заново, возрождаясь в новых поколениях, формирующих новые генерации спор, многократно повторяющиеся в течение вегетационного периода. Наибольшего развития пероноспоровые грибы достигают в ранневесенний, весенний и весеннелетний периоды. В это время года, по существу, во всех зонах обитания нарастает численность видов и частота их появления. Некоторый подъем развития пероноспоровых грибов происходит и осенью, что связано с выпадением обильных осадков и смягчением температурного режима.ОСНОВНЫЕ БОЛЕЗНИ ДАННОГО ТИПА ЭТО-Пероноспора табака, Пероноспора шпината,ПРЕНОСПОРА ХМЕЛЯ,КАРТОФЕЛЯ И ТД.
Размножение грибов осуществляется с помощью спор, а также частичек мицелия, склероциев и других элементов. Любая структура, которая может вызвать развитие молодого мицелия, называется диаспорой, или пропагулой. С учетом происхождения диаспоры подразделяют на специализированные, то есть формирующимися именно для размножения (споры), и неспециализированные (в частности, кусочки мицелия).
Размножение неспециализированными диаспорами называют вегетативным из-за того, что оно осуществляется с помощью частей или элементов вегетативных тел грибов. Относительно мицелиальных грибов следует отметить, что они размножаются исключительно посредством кусочков мицелия. Данный метод применяют для размножения многих видов грибов в культуре, к примеру шампиньонов, летнего опенка, вешенки и т.д.
Способы размножения
Споры грибов предназначены исключительно для размножения. В их структуре различают одну либо несколько клеток, которые имеют микроскопические размеры. Попадая в благоприятную среду, небольшое количество спор дают начало новому мицелию. Большинство из них гибнут, поэтому все грибы образуют огромное число спор. Так, шампиньоны дают примерно 40 млн. спор в час, трутовик – до 30 млрд.
С учетом происхождении и выполняемой функции спор в жизни грибов выделяют два способа:
1. Спорами полового размножения (мейоспорами).
2. Бесполого (митоспорами).
С помощью митоспор грибы массово расселяются на этапе вегетации при отсутствии рекомбинации наследственных свойств. При появлении новых мицелиев из мейоспор увеличивается разнообразие грибов за счет рекомбинации признаков родительских организмов. У многих грибов в жизненном цикле формируются как митоспоры, так и мейоспоры. Но у некоторых групп грибов – дейтеромицетов, объединяющих 30 тысяч видов, полностью редуцирована способность к половому размножению. Они образуют только митоспоры. Для формирования митоспор у грибов предназначены особые клетки (орган размножения) – спорангии, или веточки воздушного мицелия. Первые споры называются спорангиоспоры, а вторые, образующиеся экзогенно, – конидии.
Для формирования митоспор у грибов предназначены особые клетки (орган размножения) – спорангии, или веточки воздушного мицелия. Первые споры называются спорангиоспоры, а вторые, образующиеся экзогенно, – конидии.
Спорангиоспоры могут быть неподвижными либо со жгутиками, что дает им возможность активно передвигаться. Это так называемые зооспоры, которые перемещаются только в водной среде, а соответственно размножение возможно только в присутствии капельно-жидкой среды. Размножение зооспорами характерно для всех групп первично водных форм грибов. У наземных видов способность к их продуцированию утрачена.
У большинства грибов получило широкое распространение бесполое размножение с помощью конидий. Эти структуры формируются у высших грибов с клеточным мицелием на гифах воздушного мицелия, чаще на специальных веточках – конидиеносцах. Они мелкие и образуются в огромных количествах. Освобождение конидий осуществляется в большинстве случаев пассивно, реже возможно активное их отбрасывание, как у нигроспоры. В процессе эволюции у конидий появились множественные адаптации, способствующие их распространению.
Мейоспоры формируются у грибов внутри либо на поверхности особых клеток, которые называются сумками, асками, или базидиями. Это органы размножения грибов. Обширная группа грибов – аскомицетов, или сумчатых – характеризуется формированием сумок с мейоспорами. Базидии с мейоспорами формируются у большой группы базидиальных грибов, или базидиомицетов (около 30 тысяч видов). Освобождение базидиоспор или аскоспор происходит посредством их активного отбрасывания от базидии или аски. При слиянии двух мейоспор образуется зигота, которая при попадании в благоприятные условия дает начало новому мицелию













